Command Palette
Search for a command to run...
RNA構造予測がAlphaFold 3に匹敵!バージニア工科大学の研究チームが、進化情報に全く依存しないRNAbpFlowを提案。

2020年11月、AlphaFold 2はCASP14国際タンパク質構造予測コンペティションで一躍有名になった。タンパク質の三次元構造を実験とほぼ同等の精度で予測したAlphaFold 2は、半世紀にわたり生物学界を悩ませてきたタンパク質折り畳み問題を解決しただけでなく、その驚異的な成果により、4年後には2人の開発者がノーベル化学賞を受賞するに至った。しかし、AlphaFold 2の成功は始まりに過ぎなかった。まるで核爆弾のように、構造生物学の分野全体に火をつけ、人工知能を用いて生体高分子の空間構造を解析する研究の波を引き起こした。
AlphaFold 2がタンパク質分野で成し遂げた画期的な成果に触発され、人工知能を用いてリボ核酸(RNA)の三次元構造を予測するという課題を解決することにも大きな期待が寄せられている。しかしながら、現在の主流アルゴリズムは、実装時にいくつかの避けられない実際的なボトルネックに直面している。まず、TransformerベースのRNA予測モデルのほとんどは、多重配列アライメント(MSA)によって提供される主要な進化配列情報、または生物学的言語モデルから間接的に得られる相同配列情報に大きく依存しています。しかし、RNA塩基対合の同形構造性のため、高品質で信頼性の高いMSA結果を生成することは困難です。次に、主流の手法のほとんどは、RNAの三次元折り畳み形態を決定する中核的な要素であるRNA二次構造における塩基対合の重要な情報を十分に活用できていません。さらに、RNA分子は自然な構造的柔軟性を持ち、複数の安定した空間構造で存在します。既存のアルゴリズムは、ほとんどの場合、単一の静的な結果しか出力せず、RNAの真の動的構造セットを再構築することはできません。
前述の課題に対処するため、バージニア工科大学のデブスワプナ・バッタチャリヤ教授と彼の学生であるスミット・タラフダーは協力して、配列と塩基対の条件に基づいたSE(3)等変フローマッチングモデルであるRNAbpFlowを開発しました。このモデルは、多重配列アライメントや相同テンプレートなどの進化情報を必要とせず、RNAヌクレオチド配列と塩基対形成情報のみに基づいて、全原子RNA構造の完全なセットを生成することができる。本手法は、既存のAIベースのRNA構造予測手法のいくつかの欠点を克服するものです。複数のベンチマーク実験により、塩基対形成を制約条件として導入することで、モデルの予測精度を大幅に向上させることができることが実証されています。RNAトポロジーサンプリングと3Dコンフォメーションモデリングという2つの主要タスクにおいて、RNAbpFlowは既存の主流手法を凌駕する性能を発揮します。
RNAbpFlowとAlphaFold 3の比較テストでは、印象的なデータによって最終的な結果が明らかになったことを特筆すべきである。CASP16のブラインドテスト実験において、RNAbpFlowは14個のRNAターゲット(200ヌクレオチド以下)の大部分の全体的な折り畳みトポロジーを正確に再構築することができ、そのうち12個は適格な予測基準を満たした。対照的に、同じ実験条件下では、AlphaFold 3の予測された標的構造のうち、天然の立体構造との一致基準を満たしたのはわずか8つであった。
「RNAbpFlow:条件付きRNA 3D構造生成のための塩基対増強SE(3)フローマッチング」と題された関連研究成果は、Nature Methods誌に掲載されました。
研究のハイライト:
- 提案されたRNAbpFlowは、配列と塩基対の条件に基づくSE(3)等流体マッチングモデルであり、既存のRNA三次元構造予測方法のいくつかの欠点を克服するものです。
- 3つの主要な革新が提案された。3チャンネルの塩基対形成条件付き入力、ヌクレオシド塩基中心の特性評価、および塩基対形成に特化した補助損失(bp2Dおよびbp3D)であり、これらによって塩基対形成と三次元構造の忠実度が大幅に向上した。
- 本稿では、独立した非反復的な学習・テストセット戦略を提案し、複数の権威あるベンチマークとCASPブラインドテストを用いて検証を行う。その結果、既存の主流アルゴリズムを凌駕する総合的な性能が得られた。

用紙のアドレス:
https://www.nature.com/articles/s41592-026-03128-4
訓練データセットとテストデータセットを個別に重複排除することで、真に公平な評価が保証されます。
独自開発モデルRNAbpFlowを構築し、その評価結果の真正性と信頼性を確保するために、本研究では、内容の重複がない独立したトレーニングセットとテストセットを使用し、評価プロセス全体を通して単一のデフォルトモデルを使用するのではなく、異なるテストベンチマークに特定のモデル重みを割り当てることで、トレーニングデータが評価プロセスに混入して結果が歪むことを回避しています。
モデル開発と内部検証に関して、本研究ではRNA3DBデータセットを使用します。このデータセットは、配列レベルと構造レベルの両方で冗長性がなく、深層学習モデルのトレーニングや内部ベンチマークに非常に適しています。本研究では、2024年4月26日にタンパク質データベース(PDB)から解析されたRNA3DBデータセットを使用し、実験用の代表的なRNA配列を選択するために、原著論文で示されたトレーニング/テスト分割スキームに従いました。
データセットに高品質の天然RNA構造と正確な塩基対情報のみが含まれるようにするため、実験では多層的な品質管理フィルタリングを実施しました。これには、単一のヌクレオチドに単一の原子しかない構造やタンパク質残基が混在する構造の除去、FASTA配列と3次元構造間のミスマッチを修正して塩基対の完全性を保護するために連続した実験配列を切り詰めること、天然構造から塩基対のない配列を削除することが含まれます。RNAViewを使用して塩基対情報を抽出し、長さが20ヌクレオチド以上の連続した非対合RNA鎖をフィルタリングし、配列長を30~200ヌクレオチドに制限した後、最終的に560のRNA配列を含む純粋なトレーニングセット(Rfamファミリーに一致しない配列はデータ漏洩のリスクをさらに低減するために削除されました)と48の配列を含む純粋なテストセットが得られました。
特筆すべきは、このデータ分割部分はRNA3DBの非冗長分割に完全に準拠しており、ルートからのデータ漏洩を防ぐため、トレーニングセットとテストセットの間で配列や構造に重複がないことです。
RNAbpFlowとRNAJPの比較実験では、本研究では、RNAJP研究で使用されたものと同様の、3方向コネクターを含む22個のRNA配列のデータセットを使用し、スクリーニング処理を行った。研究者らはまず、このデータセットからマルチマー構造を除去し、残った配列を前述のクリーンなトレーニングセットと迅速に比較して冗長性を排除し、配列の重複を取り除いた。最終的に処理されたデータセットは12個の配列に絞り込まれた。
CASP15およびCASP16国際コンペティションとの互換性に関して、競技会のブラインドテスト規則を厳守し、他のアルゴリズムとの公平な比較を確保するため、本研究ではRNA3DBが提供するトレーニングセットとテストセットの分割をそのまま採用しませんでした。その代わりに、RNA3DBから完全に分離され、重複しない2つの新しいトレーニングセットを再編成し、計画しました。
CASP15では、CASP15コンペティションでテストされる最初のRNAバッチが2022年5月に公開されたばかりだったため、研究者たちは新しいトレーニングセットを設計する際に、2022年4月以前にPDBデータベースにアップロードされたRNA配列(すべてRNA3DB由来)のみを含めました。これにより、モデルのトレーニングがCASP15でテストされるRNAを完全に回避し、ブラインドテストのルールを厳守することが保証されました。最終的に、トレーニングセットは30~784ヌクレオチドの長さの731のRNA配列となりました。モデル評価に使用されたテストセットは、CASP15ベンチマークセットからの6つの天然RNAと4つの合成RNAで構成されていました。
CASP16の場合、実験で使用されたトレーニングセットは、実験的に解明された994個のPDB構造と、クロス蒸留データ拡張に使用された2,170個の高信頼性予測構造から構成され、合計3,164個のサンプルとなった。各トレーニングバッチにおけるPDB構造の数とクロス蒸留データの数の比率は約1:2.2である。
具体的には、同じ理由で、CASP16でテストされる最初のRNAバッチは2024年5月に公開されました。そのため、研究者らはRNA3DBからすべての適格データを1つのトレーニングセットに統合し、2024年4月6日以前にアップロードされたPDB構造のみを含めました。厳密なスクリーニングの後、994のRNA配列とそれに対応する測定された3次元構造が得られました。テストデータは、CASP16で現在利用可能な、実際の実験的3次元構造を持つ28のRNAから得られました。
クロス蒸留データセットの構築に使用されたデータは、主にデータ拡張の効果を調査するために、bpRNA-1m(90)データセットから取得されました。bpRNA-1m(90)データセットには、28,370個のRNA配列(およびそれに対応する二次構造)が含まれています。冗長性を除去し、配列長を30~200に制限した後、MMseqs2クラスタリングツールを使用して複数回抽出した後、最終的に得られたのは、信頼性の高いRNA構造2,170個からなるクロス蒸留データセットであった。
ストリームマッチングに基づく条件付き生成フレームワーク
RNAbpFlow は、配列と塩基対条件に基づく SE(3) 等流体マッチングモデルです (下図参照)。これは、タンパク質構造生成モデル FrameFlow を改良したもので、NuFold が提案したヌクレオチド表現法を採用し、RNA 配列の各ヌクレオチドを剛体フレームとして表現します。ヌクレオチド配列と塩基対情報を制約として使用し、反復サンプリングによって二面角を予測し、すべての原子を徐々に縮小して、真のリボースフォールド構造を持つ RNA の最終的な完全な三次元構造を作成します。つまり、RNAbpFlowの主な利点は、「入力条件」と「モデルパラダイム」という2つの点に集約されます。
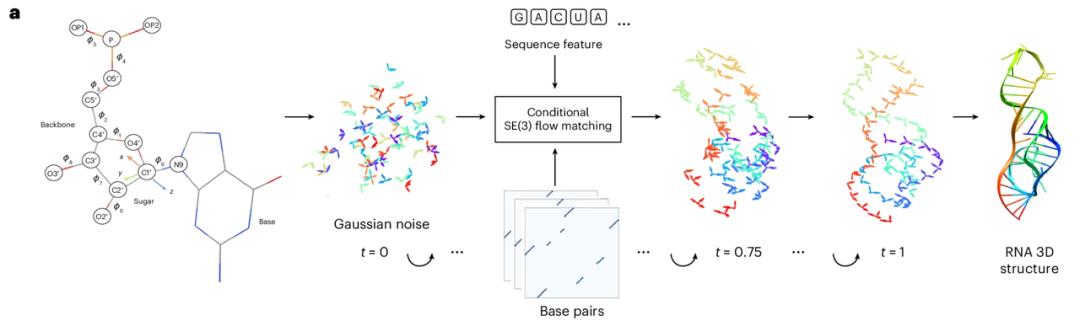
「入力条件」に関して、RNAbpFlowは、MSAや相同テンプレートの情報がなくても、配列情報と塩基対形成情報のみに基づいて、条件付きで3次元RNA構造を生成することができます。
まず、配列については、L 長の RNA 配列が入力され、ヌクレオチドはワンホットエンコーディングを使用して表現されます。つまり、4 つの要素を含むバイナリ ベクトルが 4 種類のヌクレオチド (A、U、C、および G) に対応します。次に、トレーニング フェーズでの塩基対情報処理 (これが重要なイノベーションです) では、RNAView、MC-Annotate、および DSSR の 3 つのツールを使用して、実験的に解明された自然な 3 次元構造から 2 次元のペアリング アノテーションを抽出し、これらを 3 つの独立した L × L バイナリ マトリックスとして表現します。さまざまな古典的および非古典的な塩基対の特徴を完全に捉えるために、この研究では、3 つのツールによって得られた矛盾するアノテーションを一律に修正しません。代わりに、3 つのバイナリ マトリックスを直接連結して L × L × 3 テンソルを作成し、これをノイズ除去ネットワーク構造のバイアス項への入力として使用し、3 つの独立したチャネルでペアリング特徴情報を提供します。
さらに、CASP15およびCASP16コンペティションのテストターゲットのサンプリングおよび推論シナリオでは、天然の塩基対形成の真の値がないため、実験では、IPKnot、SPOT-RNA、およびRibonanzaNetという3つのRNA 2D構造予測ツールによって出力された配列依存のペアリング予測行列を使用しました。これら3つのツールはすべて、擬似結び目識別に基づく塩基対形成予測をサポートしており、CASP15の天然RNAターゲットでのサンプリング性能に基づいて選択されました。さらに、RNAbpFlowは非常に汎用性が高く、ユーザーが入力として3セットのペアリング行列を定義することをサポートしています。カスタムペアリング行列が1セットしか利用できない場合は、それを3回コピーして、ネットワークが必要とする3チャネル2D入力形式に合わせることができます。

「モデルパラダイム」の観点から言えば、鍵となるのは「フローマッチング」です。フローマッチングは、データ分布の確率フローに一致する速度場(またはフロー場)を学習することを主な目的とする深層生成モデルの一種です。これにより、ガウス分布のような単純な分布を、高次元空間におけるターゲットに必要な複雑なデータ分布に変換します。フローマッチングは、この速度場を直接学習し、元のデータ分布を完全に破壊することなく、単純な分布からターゲット分布へ移動するサンプル点の動きを記述します。学習したベクトル場に常微分方程式を積分することで、フローマッチングはターゲットを近似するより単純な移動軌跡を生成できます。拡散モデルと比較して、フローマッチングは大規模サンプルの生成における計算速度を大幅に向上させることができます。
本研究におけるフローマッチング手法の目的は、パラメータ化されたベクトル場 Ut を学習することです。このベクトル場は、滑らかで時間とともに変化するマッピングを表し、そこから、2 つの分布、すなわち分布 p₀(T₀) (ノイズのあるフレーム) と分布 p₁(T₁) (真の参照フレーム) 間の変換関係を記述する常微分方程式が生成されます。このマッピングを学習するために、研究者らは、時刻 t におけるノイズのある真の参照フレーム Tₜ に基づいてベクトル場を予測するパラメータ化されたニューラルネットワーク Vθ(Tₜ, t) を訓練しました。このネットワークのこの部分は、FrameFlow を参考に設計されており、AlphaFold 2 の構造モジュールをバックボーンネットワークアーキテクチャとして使用しています。
具体的なトレーニング設定については、PyTorch-Lightningフレームワークを使用してモデルをトレーニングし、学習率0.0001のAdamオプティマイザを採用して実験を実施しました。分散トレーニングプロセスは、8基のNVIDIA H100 GPU上で1500回の反復処理を実行しました。
ミッションクリティカルなパフォーマンスはAlphaFold 3を凌駕する
RNAbpFlowのサンプリング性能を評価するために、本研究ではまず、粗視化分子動力学シミュレーションに基づく3次元RNA構造サンプリング手法であるRNAJPと比較した。RNAJPは、非古典的な塩基対形成、塩基スタッキング相互作用、長距離ループ間相互作用など、塩基対形成情報を明示的に考慮している。
実験結果を下図に示す。RNAbpFlowは、両方の評価指標においてRNAJPを上回った。具体的には、RNAbpFlowは平均局所距離差テストスコア(lDDTスコア)で0.66という高い値を達成したのに対し、RNAJPは0.59にとどまった。同様に、グローバルトポロジーサンプリングに関しては、RNAbpFlowは平均テンプレートモデリングスコア(TMスコア)で0.38を達成したのに対し、RNAJPは0.32であった。
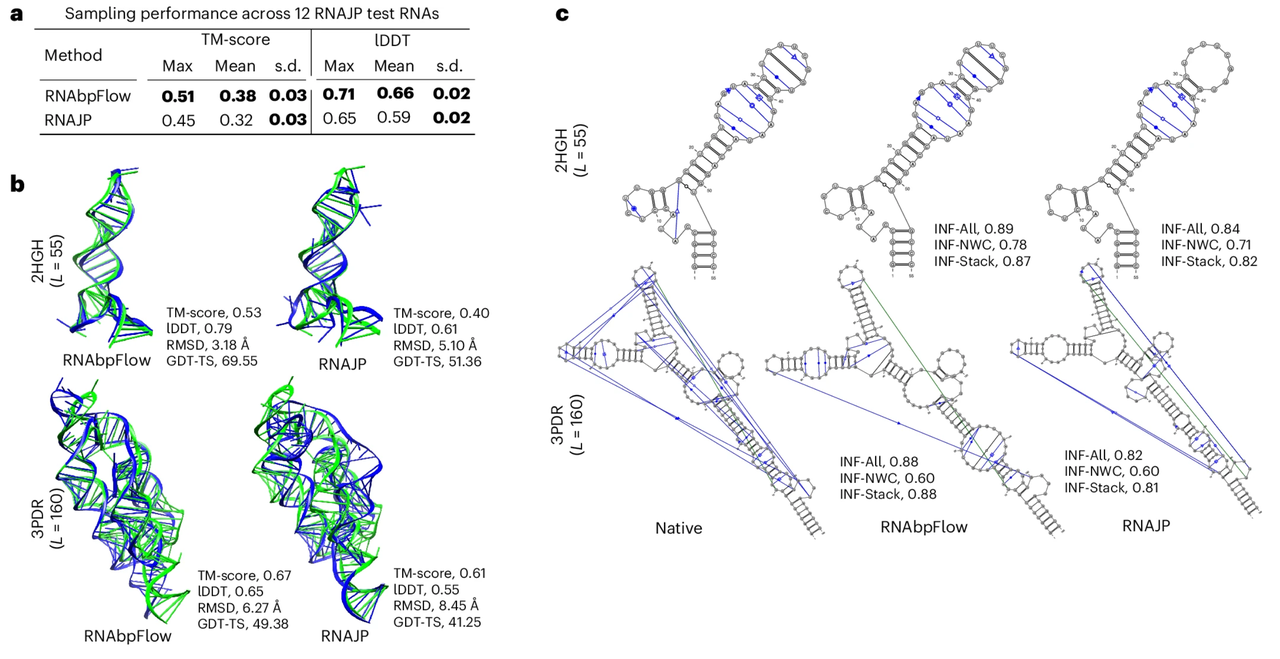
標本妥当性の評価に関しては、TMスコアに基づいて正しい折り畳み構造が決定される。RNAbpFlowは、最大で66.671 TP3TのRNA標的を検出した。lDDTスコアに基づく正しく折り畳まれた構造の検出率は251 TP3Tでした。これに対し、RNAJPの検出率はそれぞれわずか41.671 TP3Tと0 TP3Tでした。さらに、RNAbpFlowによって生成された12,000個のシミュレーションされたコンフォメーションのうち、13.41 TP3Tのコンフォメーションはテンプレートモデリングスコアが0.45より高く、9.61 TP3TのコンフォメーションはlDDTスコアが0.7より高くなりました。これに対し、RNAJPのシミュレーションされたコンフォメーションのうち、テンプレートモデリングスコアが0.45より高かったのはわずか1.731 TP3Tで、lDDTスコアが0.75より高かったコンフォメーションはありませんでした。
上記の結果は、RNAbpFlowが最適な構造のスコアリングにおいてRNAJPを上回るだけでなく、高品質なシミュレーション構造の割合もより高いことを示している。これは、グローバルなトポロジーとローカルな構成の両方を含むデュアルサンプリングタスクにおけるその効率性を強調するものである。
次に、研究チームはCASP15ブラインドテストデータセットを用いてRNA bpFlowを既存のいくつかの手法と比較した(下表参照)。その結果、…正確で実際の天然塩基対形成情報を入力すると、RNAbpFlowのモデリングおよび予測性能が大幅に向上する。平均TMスコアは0.48、全原子の二乗平均平方根偏差(RMSD)は7.77、非ワトソンクリック塩基対相互作用ネットワーク忠実度(INF-NWC)は0.62でした。アルゴリズムによって予測された塩基対のみを使用した場合、3つの指標はそれぞれ0.40、10.70、0.48でした。
それに対し、真の塩基対形成情報を提供した場合、TMスコアは201 TP3T増加し、RMSDは27.41 TP3T減少し、INF-NWCは29.21 TP3T増加し、高品質の塩基対形成情報の重要性が強調された。
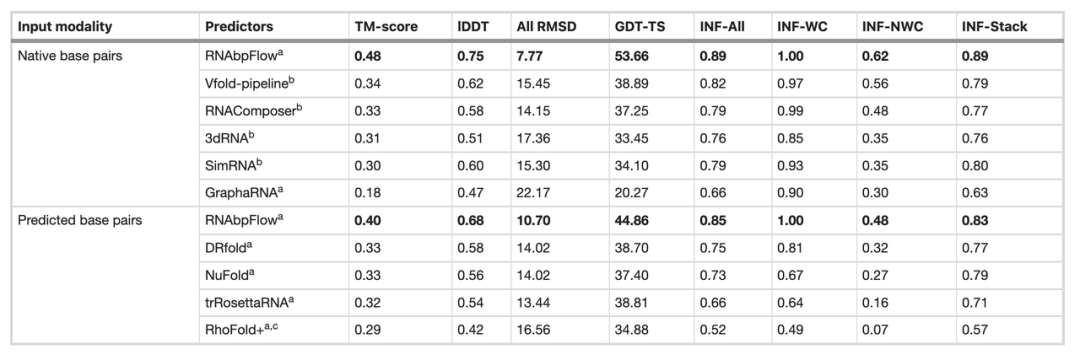
対照的に、他の手法では、実際の天然塩基対を入力した場合でも、性能向上はごくわずかしか見られません。例えば、最も性能の高いVfoldでもTMスコアは0.34にとどまり、RNAComposerの最小RMSDは14.15です。これは、RNAbpFlowがより高い適応性を持ち、深層生成モデリングにおいて正確な塩基対形成制約を効率的に利用し、AI手法の予測性能の上限を大幅に引き上げていることを示しています。
CASP16 ブラインドテストターゲットにおける RNAbpFlow のサンプリング精度を検証するため、研究者らは CASP16 コンペティションで最も優れた予測モデル 2 つと比較した。結果 (下図参照) は、14 個の予測ターゲット (≤200 nt) の平均最大 TM スコアと lDDT 指標に関して、AlphaFold 3 を含むすべてのアルゴリズムを上回る性能を示した。RNAbpFlow によって生成された構造コンフォメーションのうち、少なくとも 1 つの正しく折り畳まれたコンフォメーションが 12 (85.711 TP3T) で見つかった。これに対し、AlphaFold 3 では 8 (57.131 TP3T) のコンフォメーションでしかこれを達成できず、RNAbpFlow のより安定した性能が実証された。
しかし、200ヌクレオチドより長いRNA標的の場合、RNAbpFlowはNuFold、trRosettaRNA2、およびDRfold2よりも優れた性能を発揮するものの、AlphaFold3よりは劣ります。これは、長い配列の場合、予測される塩基対の精度が著しく低下するためです。
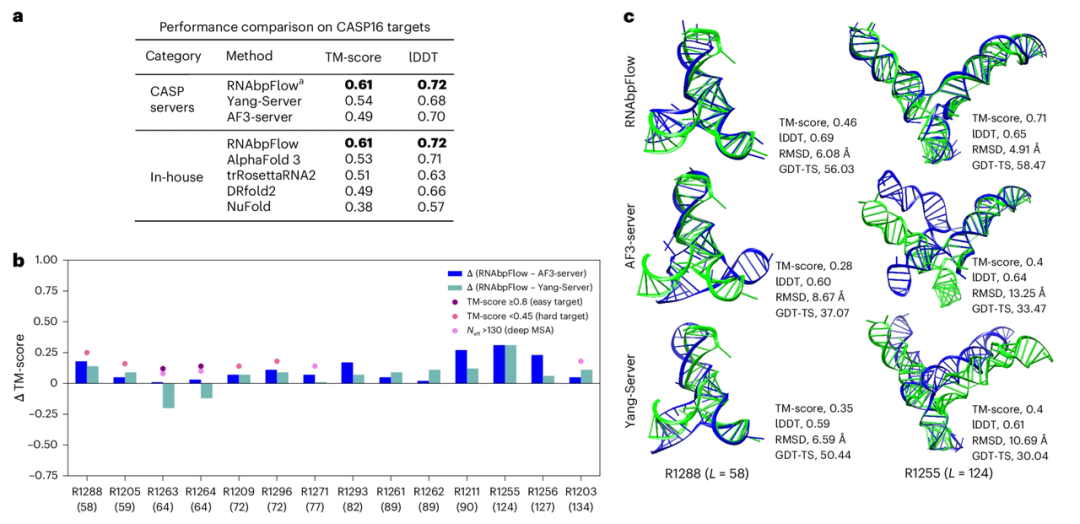
さらに、実験では、予測不可能なターゲット(TMスコア < 0.45、MSA有効相同性配列深度 ≤ 130、弱い進化シグナル)の場合、進化情報が少ないときは、塩基対形成制約に基づく構造予測モデルがRNAbpFlowを明らかに上回り、塩基対形成情報のみに基づいてRNAの三次元構造を予測できるRNAbpFlowの優位性を示した。しかし、R1263やR1264のような容易に予測可能なターゲットの場合、十分な深度を持つ多重配列アライメントデータにより、AF3-serverとYang-Serverの2つのアライメントモデルはRNAbpFlowと同等かそれ以上の性能を発揮し、両モデルが配列アライメント情報に強く依存していることが示された。
最後に書きます
要約すると、RNAbpFlowはMSAや構造相同性に制約されません。配列と塩基対形成のみを用いて、末端から末端まで直接、全原子RNAの三次元構造モデルを生成できます。高精度な原子レベルの大規模コンフォメーションセット生成技術を活用することで、RNAコンフォメーションダイナミクスの研究において、非常に有望な新たな方向性を切り開く可能性があります。