Command Palette
Search for a command to run...
ينافس التنبؤ ببنية الحمض النووي الريبي (RNA) برنامج AlphaFold 3! يقترح فريق من جامعة فرجينيا للتكنولوجيا برنامج RNAbpFlow، وهو برنامج مستقل تمامًا عن المعلومات التطورية.

في نوفمبر 2020، حقق برنامج AlphaFold 2 شهرة واسعة في مسابقة CASP14 الدولية للتنبؤ ببنية البروتينات. وبفضل دقته شبه التجريبية في التنبؤ بالبنية ثلاثية الأبعاد للبروتينات، لم يقتصر دور AlphaFold 2 على حل مشكلة طي البروتينات التي حيّرت المجتمع البيولوجي لنصف قرن، بل ساهم أيضًا في فوز مبتكريه بجائزة نوبل في الكيمياء بعد أربع سنوات بفضل نتائجه المذهلة. مع ذلك، لم يكن نجاح AlphaFold 2 سوى البداية. فقد أشعل البرنامج، كالقنبلة النووية، شرارة ثورة في مجال البيولوجيا البنيوية، مُطلقًا موجة من الأبحاث التي تستخدم الذكاء الاصطناعي لتحليل التكوين الفراغي للجزيئات الحيوية الكبيرة.
واستلهامًا من الإنجاز الرائد الذي حققه برنامج AlphaFold 2 في مجال البروتينات، تُعقد آمال كبيرة أيضًا على استخدام الذكاء الاصطناعي لحل مشكلة التنبؤ بالبنية ثلاثية الأبعاد للحمض النووي الريبي (RNA).ومع ذلك، لا تزال الخوارزميات السائدة الحالية تواجه العديد من المعوقات العملية التي لا مفر منها عند تطبيقها:أولًا، تعتمد معظم نماذج التنبؤ بالحمض النووي الريبي (RNA) القائمة على نموذج Transformer اعتمادًا كبيرًا على معلومات التسلسل التطوري السائدة المُستمدة من محاذاة التسلسلات المتعددة (MSA)، أو معلومات التسلسل المتماثل المُستقاة بشكل غير مباشر من نماذج اللغة البيولوجية. مع ذلك، فإن الطبيعة المتماثلة لتزاوج قواعد الحمض النووي الريبي تجعل من الصعب توليد نتائج محاذاة تسلسلات متعددة عالية الجودة وموثوقة. ثانيًا، تفشل معظم الطرق الشائعة في استكشاف المعلومات الأساسية لتزاوج القواعد في البنية الثانوية للحمض النووي الريبي بشكل كامل، وهو العامل الأساسي الذي يُحدد شكل الطي ثلاثي الأبعاد للحمض النووي الريبي. ثالثًا، تمتلك جزيئات الحمض النووي الريبي مرونة بنيوية طبيعية وتوجد في هيئات مكانية مستقرة متعددة. تُخرج الخوارزميات الحالية في الغالب نتائج ثابتة واحدة فقط، ولا تستطيع إعادة بناء مجموعة الهيئات الديناميكية الحقيقية للحمض النووي الريبي.
لمعالجة التحديات المذكورة أعلاه، تعاون البروفيسور ديبسوابنا بهاتاشاريا وطالبه سوميت تارافدر من جامعة فيرجينيا للتكنولوجيا لتطوير نموذج مطابقة التدفق المتغير SE(3)، RNAbpFlow، استنادًا إلى تسلسل وشروط أزواج القواعد.يمكن لهذا النموذج أن يولد مجموعة كاملة من التكوينات الذرية الكاملة للحمض النووي الريبي (RNA) بالاعتماد فقط على تسلسلات النيوكليوتيدات للحمض النووي الريبي ومعلومات اقتران القواعد، دون الحاجة إلى معلومات تطورية مثل محاذاة التسلسلات المتعددة أو القوالب المتماثلة.يعالج هذا النظام العديد من أوجه القصور في أساليب التنبؤ ببنية الحمض النووي الريبي (RNA) القائمة على الذكاء الاصطناعي. وقد أثبتت تجارب معيارية متعددة أن إدخال اقتران القواعد كقيد يمكن أن يحسن دقة تنبؤ النموذج بشكل ملحوظ. وفي المهمتين الأساسيتين المتمثلتين في أخذ عينات من بنية الحمض النووي الريبي ونمذجة التكوين ثلاثي الأبعاد، يتفوق نظام RNAbpFlow على الأساليب السائدة الحالية.
تجدر الإشارة إلى أنه في الاختبار المقارن بين RNAbpFlow و AlphaFold 3، كشفت مجموعة من البيانات الرائعة عن النتيجة النهائية للمباراة:في تجربة الاختبار العمياء CASP16، تمكن RNAbpFlow من إعادة بناء بنية الطي العالمية للغالبية العظمى من أهداف الحمض النووي الريبي الـ 14 (≤200 نيوكليوتيد) بدقة، حيث استوفى 12 منها معايير التنبؤ المؤهلة.في المقابل، وفي ظل نفس الظروف التجريبية، لم تستوفِ سوى 8 هياكل مستهدفة متوقعة من AlphaFold 3 معيار المطابقة مع التكوين الطبيعي.
تم نشر نتائج البحث ذات الصلة، بعنوان "RNAbpFlow: مطابقة تدفق SE(3) المعززة بأزواج القواعد لتوليد بنية RNA ثلاثية الأبعاد المشروطة"، في مجلة Nature Methods.
أبرز الأبحاث:
- إن نموذج RNAbpFlow المقترح هو نموذج مطابقة متساوي السوائل من نوع SE(3) يعتمد على تسلسل وشروط أزواج القواعد، والذي يعالج العديد من أوجه القصور في طرق التنبؤ بالبنية ثلاثية الأبعاد للحمض النووي الريبي الحالية.
- تم اقتراح ثلاثة ابتكارات رئيسية: إدخال مشروط لتكوين أزواج القواعد ثلاثية القنوات، وتوصيف مركز قاعدة النيوكليوزيد، وفقدان مساعد خاص بتكوين أزواج القواعد (bp2D و bp3D)، مما أدى إلى تحسين دقة تكوين أزواج القواعد والبنية ثلاثية الأبعاد بشكل كبير.
- نقترح استراتيجية مستقلة وغير متكررة لمجموعات التدريب والاختبار، ونؤكد صحة عملنا باستخدام معايير مرجعية موثوقة متعددة واختبار CASP الأعمى. ويتجاوز أداؤنا الإجمالي أداء الخوارزميات السائدة الحالية.

عنوان الورقة:
https://www.nature.com/articles/s41592-026-03128-4
يضمن إلغاء التكرار المستقل لمجموعات التدريب والاختبار تقييمًا حقيقيًا وعادلًا.
لبناء نموذج RNAbpFlow المطوّر ذاتيًا وضمان أصالة وموثوقية نتائج تقييمه، تستخدم هذه الدراسة مجموعات تدريب واختبار مستقلة بدون تكرار للمحتوى، وتخصص أوزانًا محددة للنموذج لمعايير اختبار مختلفة، بدلاً من استخدام نموذج افتراضي واحد طوال العملية، وذلك لتجنب خلط بيانات التدريب في عملية التقييم والتسبب في تشويه النتائج.
فيما يتعلق بتطوير النموذج والتحقق الداخلي منه،تستخدم هذه الدراسة مجموعة بيانات RNA3DB، وهي مجموعة بيانات ملائمة للغاية لتدريب نماذج التعلم العميق والتقييم الداخلي، إذ تتميز بعدم وجود تكرار على مستوى التسلسل والبنية. في هذه الدراسة، تم استخدام نسخة RNA3DB المُستخرجة من قاعدة بيانات البروتينات (PDB) بتاريخ 26 أبريل 2024، واتُبعت آلية تقسيم التدريب والاختبار المذكورة في الورقة البحثية الأصلية لاختيار تسلسلات RNA تمثيلية للتجارب.
لضمان احتواء مجموعة البيانات على تراكيب RNA طبيعية عالية الجودة ومعلومات دقيقة عن اقتران القواعد، خضعت التجربة لعملية ترشيح متعددة المراحل لمراقبة الجودة. شمل ذلك إزالة التراكيب التي تحتوي على ذرة واحدة فقط في نيوكليوتيد واحد، والتراكيب المختلطة ببقايا بروتينية؛ وتقصير التسلسلات التجريبية المتصلة لتصحيح عدم التطابق بين تسلسلات FASTA والتراكيب ثلاثية الأبعاد، مما يحافظ على سلامة اقتران القواعد؛ وحذف التسلسلات الخالية من اقتران القواعد من التراكيب الطبيعية. بعد استخراج معلومات اقتران القواعد باستخدام برنامج RNAView، وتصفية سلاسل RNA المتصلة غير المقترنة التي يزيد طولها عن 20 نيوكليوتيدًا، وتحديد طول التسلسل بين 30 و200 نيوكليوتيد، تم الحصول في النهاية على مجموعة تدريب نقية تحتوي على 560 تسلسل RNA (تمت إزالة التسلسلات التي لا تتطابق مع عائلة Rfam لتقليل خطر تسرب البيانات) ومجموعة اختبار نقية تحتوي على 48 تسلسلًا.
تجدر الإشارة إلى أن هذا الجزء من تقسيم البيانات يتبع تمامًا التقسيم غير المتكرر لـ RNA3DB، ولا يوجد تداخل في التسلسلات أو الهياكل بين مجموعات التدريب والاختبار، وذلك لتجنب تسرب البيانات من الجذر.
في التجربة المقارنة بين RNAbpFlow وRNAJP،استخدمت الدراسة مجموعة بيانات تضم 22 تسلسلًا من الحمض النووي الريبوزي (RNA) تحتوي على روابط ثلاثية، مشابهة لتلك المستخدمة في دراسة RNAJP، ثم أجرت عملية فحص. قام الباحثون أولًا بإزالة التراكيب متعددة الوحدات من مجموعة البيانات هذه، ثم قارنوا التسلسلات المتبقية بسرعة مع مجموعة التدريب النظيفة المذكورة سابقًا لإزالة التكرار والتخلص من تكرار التسلسلات. تم تقليص مجموعة البيانات النهائية المعالجة إلى 12 تسلسلًا.
فيما يتعلق بالتوافق مع مسابقات CASP15 و CASP16 الدولية،حرصاً على الالتزام التام بقواعد الاختبار الأعمى للمسابقة وضمان مقارنة عادلة مع الخوارزميات الأخرى، لم تعتمد الدراسة بشكل مباشر على تقسيم مجموعات التدريب والاختبار المدمج في قاعدة بيانات RNA3DB. بدلاً من ذلك، أعادت تنظيم وتخطيط مجموعتي تدريب جديدتين منفصلتين تماماً وغير متداخلتين من قاعدة بيانات RNA3DB.
بالنسبة لمسابقة CASP15، ولأن الدفعة الأولى من الحمض النووي الريبوزي (RNA) التي ستُختبر في المسابقة لم تُنشر للجمهور إلا في مايو 2022، فقد اقتصر الباحثون عند تصميم مجموعة التدريب الجديدة على تضمين تسلسلات الحمض النووي الريبوزي التي رُفعت إلى قاعدة بيانات PDB قبل أبريل 2022 (جميعها من RNA3DB). وقد ضمن هذا الإجراء تجنب تدريب النموذج تمامًا لتسلسلات الحمض النووي الريبوزي التي ستُختبر في CASP15، مع الالتزام التام بقواعد الاختبار العمياء. في النهاية، أسفرت مجموعة التدريب عن 731 تسلسلًا من الحمض النووي الريبوزي، يتراوح طولها بين 30 و784 نيوكليوتيدًا. أما مجموعة الاختبار المستخدمة لتقييم النموذج، فقد تألفت من 6 تسلسلات طبيعية و4 تسلسلات اصطناعية من مجموعة CASP15 المعيارية.
بالنسبة لـ CASP16،تتكون مجموعة التدريب المستخدمة في التجربة من 994 بنية PDB تم حلها تجريبياً و2170 بنية تنبؤ عالية الثقة تستخدم لزيادة بيانات التقطير المتقاطع، ليصبح المجموع 3164 عينة.تبلغ نسبة عدد هياكل PDB إلى عدد البيانات المقطرة بشكل متقاطع في كل دفعة تدريب حوالي 1:2.2.
وللسبب نفسه، صدرت الدفعة الأولى من الحمض النووي الريبوزي (RNA) المراد اختبارها في CASP16 في مايو 2024. ولذلك، قام الباحثون بدمج جميع البيانات المؤهلة من قاعدة بيانات RNA3DB في مجموعة تدريب واحدة، تضم فقط هياكل PDB التي تم تحميلها في أو قبل 6 أبريل 2024. وبعد عملية فحص دقيقة، تم الحصول على 994 تسلسلًا من الحمض النووي الريبوزي (RNA) وهياكلها ثلاثية الأبعاد المقاسة. وجاءت بيانات الاختبار من 28 جزيئًا من الحمض النووي الريبوزي (RNA) ذات هياكل ثلاثية الأبعاد تجريبية فعلية متوفرة حاليًا في CASP16.
استُخدمت البيانات المُستخدَمة في بناء مجموعة بيانات التقطير المتقاطع من مجموعة بيانات bpRNA-1m(90)، وذلك بشكل أساسي لاستكشاف تأثيرات زيادة البيانات. تحتوي مجموعة بيانات bpRNA-1m(90) على 28370 تسلسلًا من الحمض النووي الريبي (وبنياتها الثانوية المقابلة). بعد إزالة التكرارات وتقييد طول التسلسل إلى 30-200، وبعد عمليات استخلاص متعددة باستخدام أداة التجميع MMseqs2،وكانت النتيجة النهائية عبارة عن مجموعة بيانات التقطير المتقاطع لـ 2170 بنية RNA عالية الثقة.
إطار عمل لتوليد الشروط يعتمد على مطابقة التدفقات
يُعدّ RNAbpFlow نموذجًا للمطابقة متساوية التدفق من نوع SE(3) يعتمد على تسلسل الحمض النووي الريبوزي وشروط اقتران القواعد (كما هو موضح في الشكل أدناه). وهو مُعدّل من نموذج توليد بنية البروتين FrameFlow، ويتبنى طريقة تمثيل النيوكليوتيدات التي اقترحها NuFold، حيث يُمثّل كل نيوكليوتيد في تسلسل الحمض النووي الريبوزي كإطار صلب. وباستخدام معلومات تسلسل النيوكليوتيدات واقتران القواعد كقيود، يتنبأ النموذج بالزوايا ثنائية السطوح من خلال أخذ عينات تكرارية، مما يُقلّل تدريجيًا جميع الذرات إلى بنيتها ثلاثية الأبعاد النهائية والكاملة للحمض النووي الريبوزي مع التكوين الحقيقي لطيّ الريبوز. باختصار،يمكن تلخيص المزايا الأساسية لـ RNAbpFlow في نقطتين: "شروط الإدخال" و "نموذج النموذج".
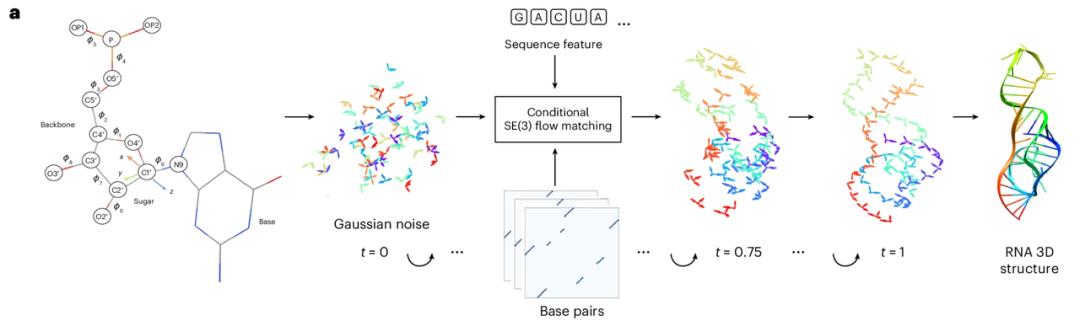
فيما يتعلق بـ "شروط الإدخال"،يمكن لبرنامج RNAbpFlow توليد هياكل RNA ثلاثية الأبعاد بشكل مشروط دون أي معلومات عن MSA أو قالب متماثل، بالاعتماد فقط على معلومات التسلسل واقتران القواعد.
أولًا، بالنسبة للتسلسل، يُدخل تسلسل RNA بطول L، وتُمثل النيوكليوتيدات باستخدام ترميز أحادي ساخن، أي أن متجهًا ثنائيًا يحتوي على أربعة عناصر يُقابل الأنواع الأربعة من النيوكليوتيدات (A، U، C، وG). ثانيًا، لمعالجة معلومات اقتران القواعد خلال مرحلة التدريب (وهذا هو الابتكار الرئيسي)، تُستخدم ثلاث أدوات - RNAView وMC-Annotate وDSSR - لاستخراج تعليقات الاقتران ثنائية الأبعاد من البنية الطبيعية ثلاثية الأبعاد التي تم تحديدها تجريبيًا، وتُمثل هذه التعليقات بثلاث مصفوفات ثنائية مستقلة بحجم L × L. ولتغطية جميع خصائص اقتران القواعد الكلاسيكية وغير الكلاسيكية، لا تُصحح الدراسة التعليقات المتناقضة التي تم الحصول عليها من الأدوات الثلاث بشكل موحد. بدلًا من ذلك، تُدمج المصفوفات الثنائية الثلاث مباشرةً في موتر بحجم L × L × 3، والذي يُستخدم كمدخل لحد الانحياز في بنية شبكة إزالة التشويش، مما يوفر معلومات خصائص الاقتران في ثلاث قنوات مستقلة.
بالإضافة إلى ذلك، في سيناريوهات أخذ العينات والاستدلال لأهداف اختبار المنافسة CASP15 وCASP16، ونظرًا لعدم وجود قيم حقيقية لأزواج القواعد الطبيعية، استخدمت التجارب مصفوفات تنبؤ الاقتران المعتمدة على التسلسل، والتي تُخرجها ثلاث أدوات للتنبؤ ببنية الحمض النووي الريبي ثنائية الأبعاد: IPKnot وSPOT-RNA وRibonanzaNet. تدعم الأدوات الثلاث جميعها التنبؤ بأزواج القواعد بناءً على تحديد العقد الكاذبة، وقد تم اختيارها بناءً على أدائها في أخذ العينات على أهداف الحمض النووي الريبي الطبيعية في CASP15. علاوة على ذلك، يتميز RNAbpFlow بتعدد استخداماته، حيث يدعم المستخدمين في تحديد ثلاث مجموعات من مصفوفات الاقتران كمدخلات. في حال توفر مجموعة واحدة فقط من مصفوفات الاقتران المخصصة، يمكن نسخها ثلاث مرات لتتوافق مع تنسيق الإدخال ثنائي الأبعاد ثلاثي القنوات المطلوب للشبكة.

من حيث "نموذج النموذج"،يكمن السر في "مطابقة التدفق". مطابقة التدفق هي نوع من نماذج التوليد العميق، هدفها الأساسي هو تعلم حقل سرعة (أو حقل تدفق) يطابق تدفق احتمالية توزيع البيانات، وبالتالي تحويل التوزيعات البسيطة، مثل التوزيعات الغاوسية، إلى توزيع البيانات المعقد المطلوب للهدف في فضاء عالي الأبعاد. تتعلم مطابقة التدفق حقل السرعة هذا مباشرةً لوصف حركة نقاط العينة التي تنتقل من توزيع بسيط إلى التوزيع المستهدف، دون إحداث تغيير جذري في توزيع البيانات الأصلي. من خلال دمج المعادلات التفاضلية العادية على حقل المتجهات المُتعلم، تستطيع مطابقة التدفق توليد مسارات انتقال أبسط لتقريب الهدف. بالمقارنة مع نماذج الانتشار، تُحسّن مطابقة التدفق بشكل ملحوظ سرعة الحساب لتوليد عينات واسعة النطاق.
تهدف طريقة مطابقة التدفق في هذه الدراسة إلى تعلم حقل متجهي مُعَلم Ut. يُمثل هذا الحقل المتجهي تحويلاً سلساً ومتغيراً مع الزمن، تُشتق منه معادلات تفاضلية عادية لوصف علاقة التحويل بين توزيعين: التوزيع p₀(T₀) (إطارات مشوشة) والتوزيع p₁(T₁) (إطارات مرجعية حقيقية). ولتعلم هذا التحويل، قام الباحثون بتدريب شبكة عصبية مُعَلمة Vθ(Tₜ, t)، والتي تتنبأ بالحقل المتجهي بناءً على الإطار المرجعي الحقيقي المشوش Tₜ عند الزمن t. صُمم هذا الجزء من الشبكة بالرجوع إلى FrameFlow، باستخدام وحدات هيكلية من AlphaFold 2 كبنية أساسية للشبكة.
فيما يتعلق بإعدادات التدريب المحددة، تم إجراء التجربة باستخدام إطار عمل PyTorch-Lightning لتدريب النموذج، باستخدام مُحسِّن Adam بمعدل تعلم قدره 0.0001. تم تشغيل عملية التدريب الموزعة على 8 وحدات معالجة رسومية NVIDIA H100 لمدة 1500 تكرار.
يتجاوز الأداء في المهام الحرجة أداء AlphaFold 3
لتقييم أداء أخذ العينات لـ RNAbpFlow، قارنت الدراسة أولاً بينه وبين RNAJP، وهي طريقة لأخذ عينات من بنية الحمض النووي الريبي ثلاثية الأبعاد تعتمد على محاكاة ديناميكيات الجزيئات الخشنة التي تأخذ في الاعتبار بشكل صريح معلومات اقتران القواعد، وتغطي اقتران القواعد غير الكلاسيكي، وتفاعلات تكديس القواعد، وتفاعلات الحلقة-الحلقة طويلة المدى.
تظهر نتائج التجربة في الشكل أدناه.تفوق برنامج RNAbpFlow على برنامج RNAJP في كلا مقياسي التقييم.على وجه التحديد، حقق برنامج RNAbpFlow متوسطًا عاليًا في اختبار فرق المسافة المحلية (lDDT score) بلغ 0.66، بينما حقق برنامج RNAJP 0.59 فقط. وبالمثل، فيما يتعلق بأخذ عينات الطوبولوجيا العالمية، حقق برنامج RNAbpFlow متوسطًا في نمذجة القوالب (TM-score) بلغ 0.38، بينما حقق برنامج RNAJP 0.32.
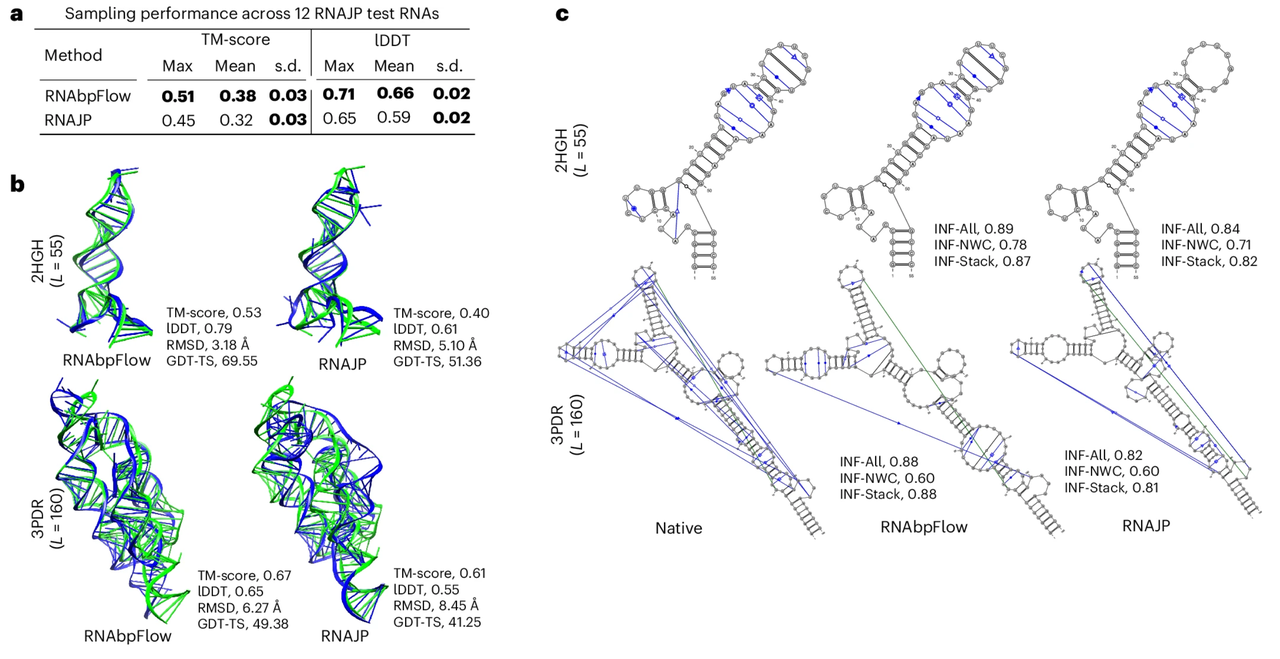
فيما يتعلق بتقييم صحة أخذ العينات، يتم تحديد بنية الطي الصحيحة بناءً على درجة TM.كشف برنامج RNAbpFlow عن أهداف الحمض النووي الريبي (RNA) التي تصل إلى 66.671 TP3T.بلغ معدل اكتشاف البنى المطوية بشكل صحيح بناءً على درجات lDDT 251 TP3T؛ في المقابل، كانت المعدلات لـ RNAJP 41.671 TP3T و0 TP3T على التوالي. علاوة على ذلك، من بين جميع التكوينات المحاكاة البالغ عددها 12000 التي أنشأها RNAbpFlow، كان لدى 13.41 تكوينًا من نوع TP3T درجات نمذجة قالب أعلى من 0.45، وكان لدى 9.61 تكوينًا من نوع TP3T درجات lDDT أعلى من 0.7. في المقابل، من بين التكوينات المحاكاة لـ RNAJP، كان لدى 1.731 تكوينًا فقط من نوع TP3T درجات نمذجة قالب أعلى من 0.45، ولم يكن لدى أي تكوين درجات lDDT أعلى من 0.75.
تُظهر النتائج المذكورة أعلاه أن RNAbpFlow لا يتفوق فقط على RNAJP في تسجيل الهياكل المثلى، ولكنه يحقق أيضًا نسبة أعلى من التكوينات المحاكاة عالية الجودة.وهذا يسلط الضوء على كفاءتها في مهام أخذ العينات المزدوجة للطوبولوجيا العالمية والتكوين المحلي.
بعد ذلك، قارن فريق البحث برنامج RNA bpFlow مع عدة طرق موجودة على مجموعة بيانات اختبار CASP15 العمياء (كما هو موضح في الجدول أدناه). وتُظهر النتائج أن...عند إدخال معلومات حقيقية ودقيقة عن اقتران القواعد الطبيعية، يتم تحسين أداء النمذجة والتنبؤ لبرنامج RNAbpFlow بشكل كبير.بلغ متوسط درجة TM 0.48، وكان متوسط الانحراف التربيعي لجميع الذرات (RMSD) 7.77، وكانت دقة شبكة تفاعل أزواج القواعد غير واتسون-كريك (INF-NWC) 0.62؛ وعندما تم استخدام أزواج القواعد المتوقعة بواسطة الخوارزمية فقط، كانت المؤشرات الثلاثة 0.40 و10.70 و0.48 على التوالي.
بالمقارنة، عند تقديم معلومات الاقتران القاعدي الحقيقية، زادت درجة TM بمقدار 201 TP3T، وانخفض RMSD بمقدار 27.41 TP3T، وزاد INF-NWC بمقدار 29.21 TP3T، مما يسلط الضوء على أهمية معلومات الاقتران القاعدي عالية الجودة.
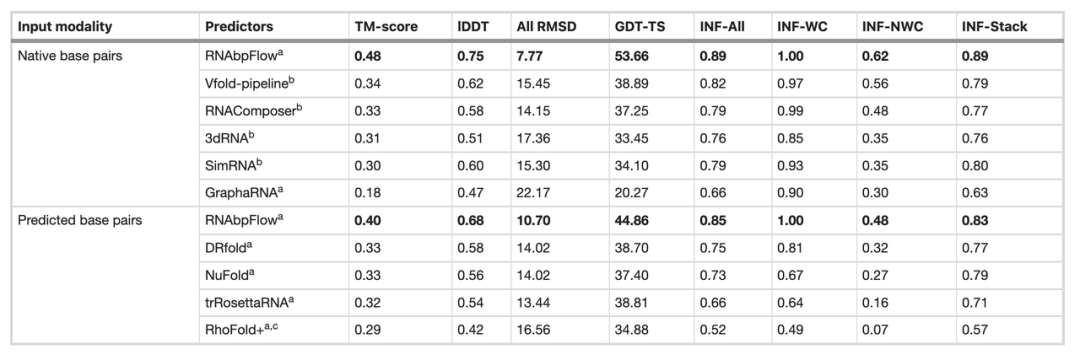
في المقابل، تُظهر طرق أخرى تحسناً محدوداً للغاية في الأداء حتى عند إدخال أزواج قواعد طبيعية حقيقية. على سبيل المثال، لا يحقق Vfold، وهو الطريقة الأفضل أداءً، سوى درجة TM تبلغ 0.34، بينما يبلغ الحد الأدنى لمتوسط الانحراف التربيعي (RMSD) لـ RNAComposer 14.15. يُظهر هذا أن RNAbpFlow يتمتع بقدرة أكبر على التكيف، حيث يستغل بكفاءة قيود اقتران القواعد الدقيقة في النمذجة التوليدية العميقة، ويرفع بشكل ملحوظ الحد الأعلى للأداء التنبؤي لأساليب الذكاء الاصطناعي.
للتحقق من دقة أخذ العينات لنموذج RNAbpFlow على أهداف اختبار CASP16 العمياء، قارن الباحثون أداءه مع أفضل نموذجين للتنبؤ في مسابقة CASP16. تُظهر النتائج (كما هو موضح في الشكل أدناه) تفوق RNAbpFlow على جميع الخوارزميات، بما في ذلك AlphaFold 3، من حيث متوسط أعلى قيمة لـ TM-score وlDDT لـ 14 هدفًا متوقعًا (≤200 نيوكليوتيد). من بين التكوينات الهيكلية التي أنشأها RNAbpFlow، وُجد تكوين واحد على الأقل مطوي بشكل صحيح في 12 منها (85.711 TP3T)؛ في المقابل، حقق AlphaFold 3 ذلك في 8 تكوينات فقط (57.131 TP3T)، مما يدل على الأداء الأكثر استقرارًا لـ RNAbpFlow.
مع ذلك، بالنسبة لأهداف الحمض النووي الريبوزي (RNA) التي يزيد طولها عن 200 نيوكليوتيد، لا يزال برنامج RNAbpFlow يتفوق على برامج NuFold وtrRosettaRNA2 وDRfold2، ولكنه أقل كفاءة من برنامج AlphaFold3. ويعود ذلك إلى انخفاض دقة التنبؤ بتطابق القواعد في التسلسلات الطويلة بشكل ملحوظ.
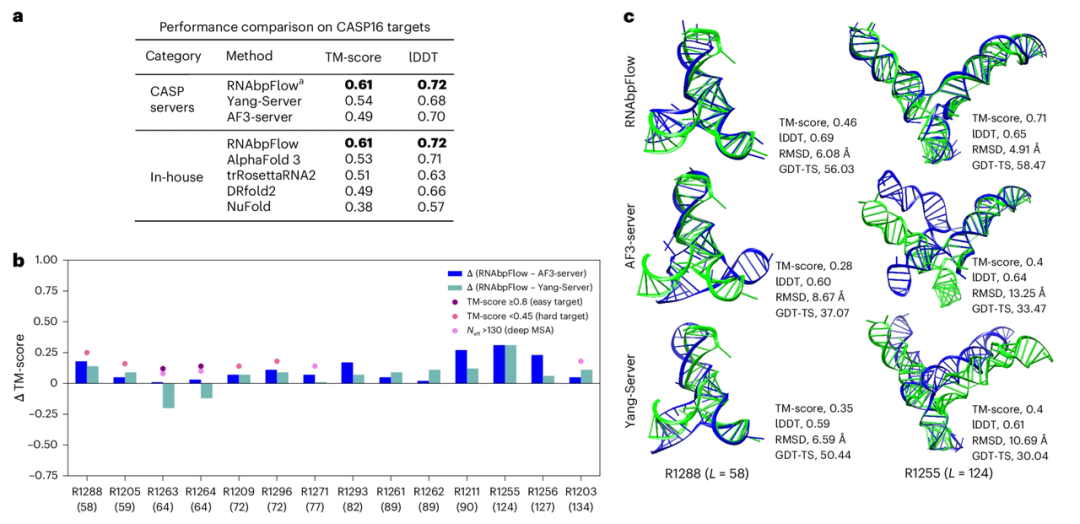
علاوة على ذلك، أظهرت التجارب أنه بالنسبة للأهداف غير المتوقعة (درجة TM < 0.45؛ عمق تسلسل التماثل الفعال لـ MSA ≤ 130؛ إشارة تطورية ضعيفة)، تفوق نموذج التنبؤ بالبنية القائم على قيود اقتران القواعد بشكل واضح على RNAbpFlow عندما كانت المعلومات التطورية شحيحة، مما يُبرز تفوق RNAbpFlow، الذي يمكنه التنبؤ بالبنية ثلاثية الأبعاد للحمض النووي الريبي (RNA) بالاعتماد فقط على معلومات اقتران القواعد. مع ذلك، عند مواجهة أهداف يسهل التنبؤ بها مثل R1263 وR1264، مكّنت بيانات محاذاة التسلسل المتعددة العميقة الكافية نموذجي المحاذاة، AF3-server وYang-Server، من أن يكونا على قدم المساواة مع RNAbpFlow أو حتى أفضل منه، مما يُظهر اعتماد كلا النموذجين بشكل كبير على معلومات محاذاة التسلسل.
الكلمات الأخيرة
باختصار، لا يقتصر برنامج RNAbpFlow على تسلسل الأحماض الأمينية المتعددة أو التماثل البنيوي. فهو قادر على توليد نماذج ثلاثية الأبعاد لبنية الحمض النووي الريبي (RNA) كاملة الذرات من البداية إلى النهاية باستخدام التسلسل واقتران القواعد فقط. وبفضل تقنية توليد مجموعات التشكيل واسعة النطاق عالية الدقة على المستوى الذري، قد يفتح هذا البرنامج آفاقًا جديدة واعدة لدراسة ديناميكيات تشكيل الحمض النووي الريبي.








