Command Palette
Search for a command to run...
触媒能力3.5倍アップ!中国科学院のチームは、拡散モデルに基づいて、P450 酵素の新規設計法である P450Diffusion を開発しました。

シトクロム P450 酵素は、ほぼすべての生物に遍在し、生命の成長と発達におけるさまざまな代謝プロセスで重要な役割を果たしています。自然界で最も多用途な生体触媒として、P450 酵素は、95% で報告されている酸化還元反応を触媒するだけでなく、温和な条件下で不活性な炭素-水素結合を選択的に酸化することもできます。産業用途では「万能触媒」として知られています。
現在、より優れた性能を持つ新しい P450 酵素を設計するために指向進化が広く使用されていますが、従来の方法では通常、複数回のランダム変異誘発とハイスループットスクリーニングが必要なため、実際の実験やコンピューターシミュレーションによる計算を網羅的に実行することは困難です。根底にあるタンパク質空間を探索します。
ディープラーニングはタンパク質の構造予測において目覚ましい成果を上げていますが、理想的な機能設計は依然として大きな課題です。タンパク質の機能設計を行う際に、十分な高品質の機能データを収集し、複雑なモデルをトレーニングして目的の配列を作成することは困難です。したがって、機能知識主導型の技術と強力な深層学習モデルを組み合わせて、天然のタンパク質配列空間を拡張します。は、新規 P450 酵素を設計するための適切なアプローチである可能性があります。
最近、中国科学院天津産業生物技術研究所のJiang Huifeng氏やCheng Jian氏らの研究者らは、P450酵素フラボン6-ヒドロキシラーゼ(F6H)のポケット設計原理を分析した。拡散モデルとポケット設計原理に基づいた P450 酵素の新規設計法である P450Diffusion を開発しました。関連する研究は、「拡散モデルにおける触媒ポケットを制約することによるチトクローム P450 酵素設計」というタイトルで Research 誌に掲載されました。
この研究では、P450Diffusion に基づいて、天然の P450 酵素よりも優れた活性と高い安定性を備えた新しい酵素を生成しました。この新しい酵素の触媒能力は、天然のフラボノイド 6 位水酸化酵素と比較して 1.3 ~ 3.5 倍増加しました。
研究のハイライト:
※本研究は、P450酵素の進化における新たな機能の起源メカニズムを解析し、P450酵素の基質結合における「3点固定原理」を提案するものです。
※P450Diffusionにより生成される新たな酵素の触媒能力は1.3~3.5倍に増加
* この研究は、深層学習拡散モデルの枠組みの下で新しい機能性 P450 酵素を設計するための新しいアイデアを提供します。将来的には、この手法はバイオエンジニアリングや産業触媒分野での役割を果たし、新たな酵素の開発と応用を促進することが期待されます。

用紙のアドレス:
https://spj.science.org/doi/10.34133/research.0413
オープンソース プロジェクト「awesome-ai4s」は、100 を超える AI4S 論文の解釈をまとめ、大規模なデータ セットとツールを提供します。
https://github.com/hyperai/awesome-ai4s
データセット: データセットを収集してコード化する
P450Diffusion を構築するために、研究者らは、公開されている P450 酵素データベースおよび公共データベースからすべての潜在的な P450 酵素をスクリーニングおよび分析し、560 を超える長さの配列を除外し、トレーニング データセットとして 226,509 個の配列を取得しました。
次に、研究者らはトレーニング データ セットをエンコードし、タンパク質配列の各アミノ酸を 8 次元ベクトルとしてエンコードし、タンパク質配列の各バッチを 64×1×560×8 ベクトルとしてエンコードしました (64 はバッチ サイズ)。 . はトレーニング データのサンプル数に等しく、1 はチャネル サイズを表し、8 はタンパク質配列の各アミノ酸の VHSE8 エンコード ベクトルを表します。
タンパク質配列が 560 より短い場合、研究者らは長さが 560 に達するまでギャップを追加しました。この場合、ギャップのエンコードとして 8 つのゼロのベクトルが割り当てられます。
モデルアーキテクチャ: P450 酵素 de novo 設計法 P450Diffusion
研究者らはエリゲロンのフラボノイド 6-ヒドロキシラーゼ (CYP706X1) を例に挙げました。この酵素は CYP706X サブファミリーに属し、スクテラリン生合成経路でアピゲニンをスクテラリンに変換します。
初め、研究者らは、祖先配列の再構築、復帰突然変異実験、漸進的順方向蓄積および結晶学的解析を通じて、P450酵素遺伝子機能の革新に関与する触媒ポケットを構成する基礎残基を特定した。第二に、研究者らは、詳細な構造解析を通じて、機能的に革新的な触媒ポケットの設計原理を解明しました。やっと、触媒ポケットの設計原理と、画像生成に優れたノイズ除去拡散確率モデルを組み合わせて、人工 P450 酵素生成モデル P450Diffusion を設計しました。
ステップ 1: 触媒ポケットを構成し、P450 酵素遺伝子機能の革新に関与する基礎残基を決定します。
同定されたCYP706ファミリーのP450酵素のうち、CYP706XサブファミリーのP450酵素のみがフラボノイド基質を触媒することができ、これはP450酵素のフラボノイド6-ヒドロキシラーゼ(F6H)の機能がCYP706Xサブファミリーの祖先に新たに発明された可能性を示唆している。 。
F6H機能を持つ触媒ポケットの形成の分子機構を解明するために、研究者らは、非機能性ancXY(CYP706XおよびCYP706Yサブファミリーの共通祖先)と機能性ancX(共通祖先)の間の残基を分析することを提案した。 CYP706X サブファミリーの触媒ポケットの組成の変化。活性中心から 8 Å 以内では、48 残基のうち 16 個が異なります。
以下の図 A に示すように、ancX と ancXY の残基はそれぞれシアンとマゼンタに染色され、16 残基すべてが ancX の対応する残基で置換されると、変異体 (ancXY-16 と呼ばれます) は、図に示すように F6H 機能を獲得しました。下の図B。
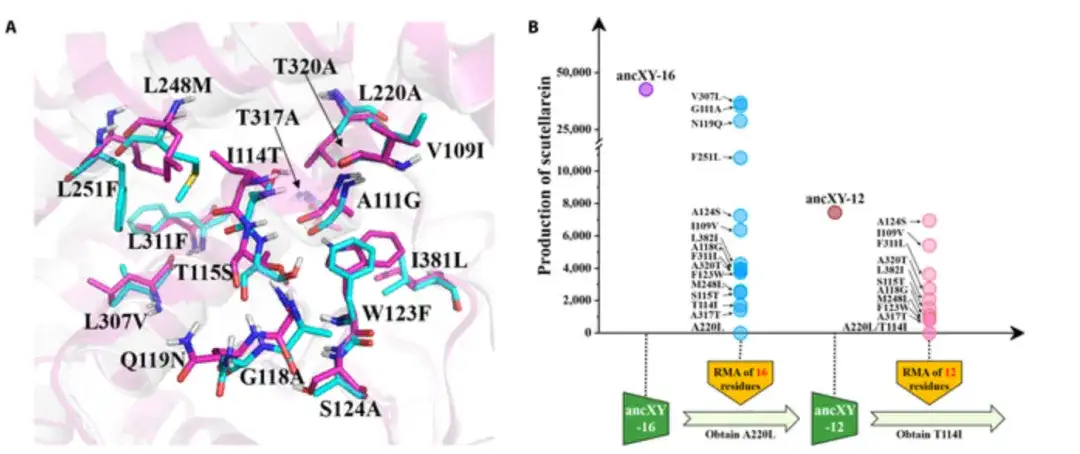
触媒ポケット内の残基の位置が三次元空間で異なり、すべての残基が基質の認識と結合に大きく寄与するわけではないことを考慮して、研究者らは復帰突然変異アッセイ (RMA: ancXY の各残基を変換することによる) の使用を試みました。 -16 触媒ポケット (変異の影響を評価するために塩基をその祖先型に戻す) を行い、触媒ポケット内の最初の残基を同定します。基礎となる残基をより迅速に特定するために、研究者らはまた、プログレッシブフォワード蓄積(PFA)戦略を使用して、変異体がF6H機能を獲得するまで重要な変異をancXYに徐々に追加した。
最後に、実験により、5つのアミノ酸変異(L220A/I114T/T317A/W123F/L248M)が、ancXYからancXへのF6Hの機能革新プロセスにおいて創始者の役割(基礎残基)を果たしていることが判明した。
ステップ 2: 機能的に革新的な触媒ポケットの設計原理を解明する。
ancXY-5のアピゲニン結合モデルの詳細な分析を通じて、研究者らは、機能革新に関与する5つの基礎残基の潜在的なメカニズムをさらに解読しました。 5 つの基礎残基の変異に基づいて、触媒ポケットは「3 点固定」原理に従っているように見えます。
「三点固定」とは、アピゲニン分子内の 3 つのハブとの重要な相互作用を指します。アピゲニン分子の 4'-OH (最初のハブ) は T114 によってもたらされる水素結合によって固定され、アピゲニンの「B」環 (2 番目のハブ) は F123 と M248 の π スタッキング相互作用によって固定されます。セロリ 分子の 7-OH (3 番目のハブ) は、水素結合によって CpdI の鉄-酸素基に固定されています。
このモデルは、基質アピゲニンを準反応性立体配座 (NAC) に維持し、アピゲニン反応部位と CpdI 鉄酸素基の間の相対配向を適切な距離と角度 (3.6 Å および 155°) で維持します。 -触媒プロセス中のアピゲニンの水酸化反応。
研究者らは、「3点固定」がF6Hの天然機能革新のための触媒ポケット設計原理として使用できることを提案し、これはまた、所望の機能を有するP450酵素を設計するための新しいアイデアを提供する。
ステップ 3: 触媒ポケットの設計原理と画像生成に優れたノイズ除去拡散確率モデルを組み合わせて、人工 P450 酵素生成モデル P450Diffusion を設計します。
研究者らは、以下の図に示すように、拡散モデルと F6H 触媒ポケットの設計原理を組み合わせて、望ましい機能を持つ P450 酵素をゼロから設計しました。新しい P450 酵素の設計プロセスには、P450 拡散モデルの構築、配列生成、スクリーニング、および実験的検証が含まれます。
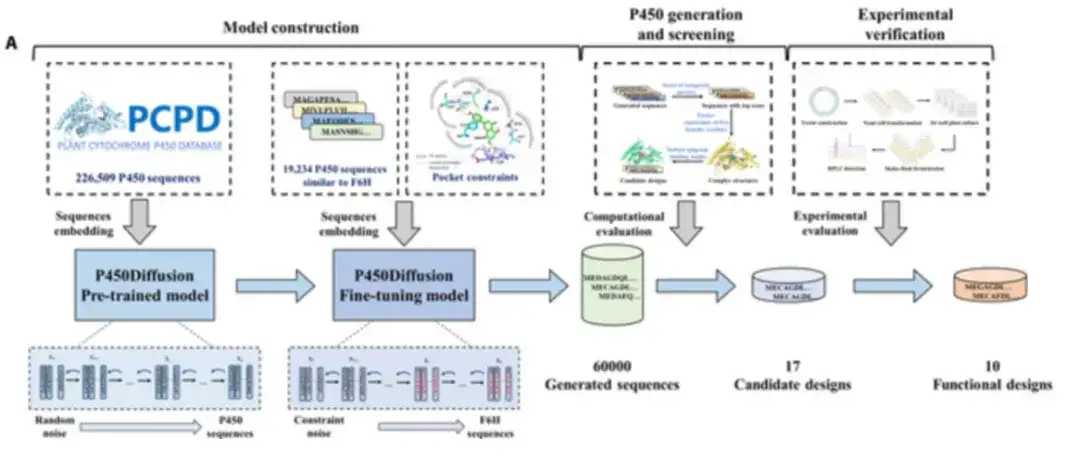
P450Diffusion には主に、事前トレーニング モデルと微調整モデルの 2 つのモデルが含まれています。
初め、事前トレーニングされた P450 配列拡散モデルをトレーニングするために、226,509 個の天然 P450 酵素配列が収集されました。
事前トレーニングされたモデルは 2 つのサブプロセスで構成されます。順拡散サブプロセスは、ランダム ノイズになるまで P450 酵素配列の表現にガウス ノイズを徐々に追加します。逆生成サブプロセスは、ランダム ノイズから始まります。そして、P450酵素配列の表現を段階的にノイズ除去して、新しいP450配列を生成する。 150,547 ラウンドのトレーニングの後、事前トレーニングされた拡散モデルは、20% から 50% までの範囲の自然シーケンスとの類似性を持つさまざまなシーケンスを生成できます。
第二に、事前トレーニングされた拡散モデルは、CYP706X サブファミリーと顕著な類似性を持つ 19,202 個の P450 酵素配列を使用して微調整され、生成された配列が F6H と同様の構造バックボーンを持つことを確認しました。
さらに、5 つの基本残基 T114、F123、A220、M248、および A317 は、「3 点固定」設計原理が de novo 生成されたシーケンスで確実に再現できるように制約され、トレーニング セットの微調整が組み合わされました。制約生成を伴うモデルは、微調整拡散モデルと呼ばれます。
続いて、実験的検証の成功率を向上させるために、研究者らは 3 つの基準を使用して 60,000 個の生成された配列の仮想スクリーニングを実行しました。生成された配列の品質を評価するための包括的な指標の計算されたスコア、5 つの基礎残基の 3 次元ポケット制約、とアピゲニンの組み合わせモデルの堅牢性。
バーチャル上映後、研究者らは、さらなる探索のために、17 の有望な非天然 P450 酵素を慎重に選択しました。
研究結果:触媒能力が1.3~3.5倍に向上
研究者らは、P450Diffusion によって生成された配列が本物の P450 酵素であり、F6H 機能を実行するかどうかを実験的にテストしました。
仮想スクリーニングの後、研究者らは選択された 17 のデザインを合成し、酵母発現システムで発現させました。 CYP706X1と比較すると、これらのデザインは、70% から 87% までの配列同一性を示します。新規触媒としての可能性を強調しています。
基質としてアピゲニンを供給して組換え酵母を 4 日間培養し、HPLC 分析を行うことにより、研究者らは、以下の図に示すように、顕著な F6H 活性を持つ 10 個のデザインを発見しました。
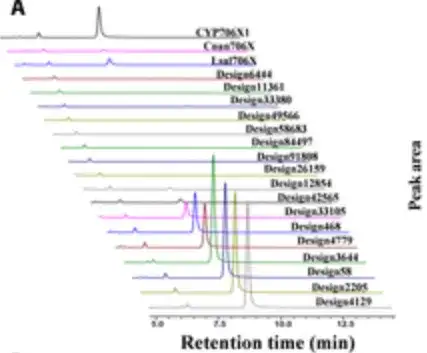
驚くべきことに、6 つの設計では、CYP706X1 と比較して、黄色の花オレアンドリン生成の触媒能力が 1.3 ~ 3.5 倍増加したことが示されました。以下の図の右側の 6 本のバーで示されているように、残りの 4 つの活性デザインも他の天然 F6H 酵素 (つまり、Cnan706X および Lsal706X) と同等の活性を示しました。
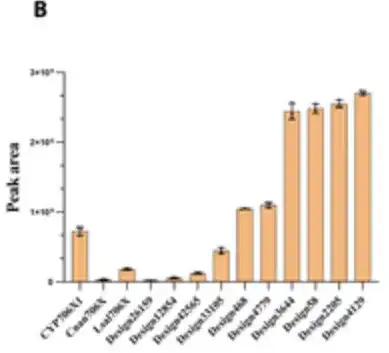
結果は、P450Diffusion が F6H 触媒ポケットの基本設計原理を捕捉し、F6H 活性を持つ P450 酵素配列を効果的に生成できるだけでなく、また、P450 酵素配列空間から天然配列よりも優れた P450 酵素をスクリーニングすることも可能です。
データ駆動型の機械学習は酵素の進化を加速するのに役立ちます
自然界に存在する酵素にはさまざまな機能があり、工業生産や学術研究に利用されていますが、その多くは用途のニーズに十分に応えられず、その特性を改良する必要があります。酵素工学のタスク。
で、指向進化法は、自然界の進化過程をシミュレートすることにより、酵素の進化速度を高めることができます。それは酵素分子の変換のための重要な技術となっています。指向性進化は生体触媒作用や創薬において重要な役割を果たしていますが、突然変異のランダム性により多数の突然変異体が生成されるため、実験的スクリーニング能力は大きな課題に直面しています。近年、人工知能やビッグデータ処理などの新興技術が生体触媒分野の重要な研究手法に発展しました。その中でも、機械学習はデータ駆動型の方法で配列/構造と酵素の機能のマッピングを取得し、酵素工学の効率を向上させるのに役立ちます。
酵素をコードする遺伝子は容易に同定できますが、大部分(99% を超える)では、酵素機能の実験による特徴付け、つまり、特定の酵素が何に変換されるか、開始分子を特定の分子に変換するため、シンテターゼの正確な機能は不明です。末端分子の作成には非常に時間がかかります。
この課題に応えて、デュッセルドルフ大学 (HHU) の研究者は、酵素と基質のペアを予測するための一般的な機械学習モデル ESP を開発しました。独立した多様なテストデータで精度は 91% を超えます。 ESP は、トレーニング データに含まれるさまざまな酵素やさまざまな代謝物にうまく適用でき、十分に研究された個々の酵素ファミリー向けに設計されたモデルよりも優れたパフォーマンスを発揮します。
この研究は、「機械学習と深層学習に基づいて酵素の小分子基質を予測する一般モデル」というタイトルで、2023年5月にNature Communications誌に掲載された。
論文リンク:
https://www.nature.com/articles/s41467-023-38347-2
新しい酵素の設計は刺激的である一方で、酵素触媒作用の複雑さによる課題も抱えています。無細胞酵素エンジニアリング会社 Enzymit の研究者は、酵素設計の新しい戦略である CoSaNN (Conformational Sampling using Neural Networks) を導入しました。深層学習の進歩を構造予測とシーケンス最適化に活用します。酵素の立体構造を制御することにより、研究者は単純な突然変異誘発の範囲を超えて化学空間を拡大できます。
さらに、チームは SolvIT をさらに開発しました。これは、大腸菌におけるタンパク質の溶解度を予測するために訓練されたグラフ ニューラル ネットワークです。高度に発現された酵素を生成するための最適化の追加層として機能します。この方法を通じて、研究者らは優れた発現レベルを持つ新規酵素を設計した。その中で、54%の設計は大腸菌で発現し、熱安定性が向上し、30%以上の設計は鋳型酵素よりも高いTmを示した。
この研究は「ディープラーニングを使用した誘導適合酵素のコンテキスト依存設計は、発現が良く、熱的に安定で活性な酵素を生成する」と題され、2023年8月にbioRxivプレプリントプラットフォームで公開された。
論文リンク:
https://www.biorxiv.org/content/10.1101/2023.07.27.550799v3

データによると、2023 年の世界市場規模は工業用酵素だけでも 74 億米ドルに相当します。将来的には、人工知能の力を利用してタンパク質の組成と進化に関する特徴的な情報を学習することで、研究者は有益な効果をもたらす変異の予測、タンパク質の安定性の最適化、触媒活性の改善、酵素工学のさまざまな問題を解決できるようになるでしょう。もっと... . これにより、バイオ製造のコストがさらに削減され、より高い商品価値がもたらされます。
参考文献:
1.https://spj.science.org/doi/10.34133/research.0413
2.https://www.cas.cn/syky/202407/t20240718_5026250.shtml
3.https://biotech.aiijournal.com/CN/10.13560/j.cnki.biotech.bull.1985.2022-0724
4.https://www.jiqizhixin.com/articles/2023-06-25-12
5.https://www.jiqizhixin.com/articles








